
大刺鳅Y染色体的端粒-端粒组装揭示了年轻性染色体对的起源
2021年7月12日,福建省淡水水产研究所薛凌展团队联合福建师范大学、华中农业大学和维也纳大学等单位,在Genome Biology在线发表题为Telomere-to-telomere assembly of a fish Y chromosome reveals the origin of a young sex chromosome pair的论文,从染色体层面解析了大刺鳅基因组,对性染色体的起源及重组抑制进行了相关研究,构建了鱼类Y染色体完整图谱,提出动物性染色体近着丝粒起源的假说。

性染色体的起源需要建立 proto-sex chromosomes 之间的重组抑制机制。在许多鱼类中,性染色体对是同态的,并且最近起源的,这为研究重组抑制如何以及为什么在性染色体分化的初始阶段进化提供了物种基础,但这需要X和Y(或Z和W)染色体的精确序列组装,如果他们最近出现分化的,这可能会很困难。
研究人员构建了一个染色体水平的单倍体型的大刺鳅(Mastacembelus armatus)的基因组。其二倍体组装几乎没有gap,在大多数染色体上,解决了中着丝粒和亚中着丝粒的异染色质序列。Y染色体包括其高度重复的短臂,没有gap。利用重测序数据,确定了一个约7Mb的完全性连锁区域(SLR),横跨性染色体的着丝粒区域,几乎完全嵌入在着丝粒周围的异染色质中。X和Y染色体上的SLR在序列和基因内容上几乎相同,但都是重复的序列和异染色质,几乎没有重组发生。进一步确定SLR中的一个含HMG域的基因HMGN6是一个候选的性别决定基因,在睾丸发育开始时表达。
背景
- 性染色体是从一对普通常染色体进化而来的。
- 理论研究表明,当性别拮抗多态性建立在一个性别决定位点附近时,性连锁区(SLR)的重组抑制被选择。
- 在真兽类哺乳动物和新颌总目的鸟类中,性染色体(Y或W)是基因贫乏、高度重复的以及含有异染色质。这种退化过程似乎是重组抑制的必然结果。
- 在其他类群中,包括非鸟类的爬行类、两栖类和鱼类,性染色体通常是同态的。一些同态性染色体的起源较近,因此是研究性染色体分化早期阶段的极好模型。
- 组装Y(或W)染色体序列时,与X(或Z)的差异很小,对于组装来说十分困难。这部分是因为Y染色体连锁区域可能是高度重复的以及富含异染色质的,使得很难通过短读长方法进行拼接。
- 年轻性染色体对的单倍型分解组装有助于解决性染色体分化的早期事件,解决诸如重组为什么和如何被抑制,以及什么特性使一个区域易于成为性连锁区域等问题。
- 大刺鳅属于合鳃目,单倍体数为24,性别发生由雌转雄的变化。重要的是,最近的一项研究发现了两个male特异性分子标记,这意味着XY性染色体的存在,尽管细胞遗传学上所有的染色体对都是同态的。这使大刺鳅成为研究性染色体起源和性染色体表达与性别变化之间相互作用的一个极好的模型。
结果
单倍型解析的染色体水平组装
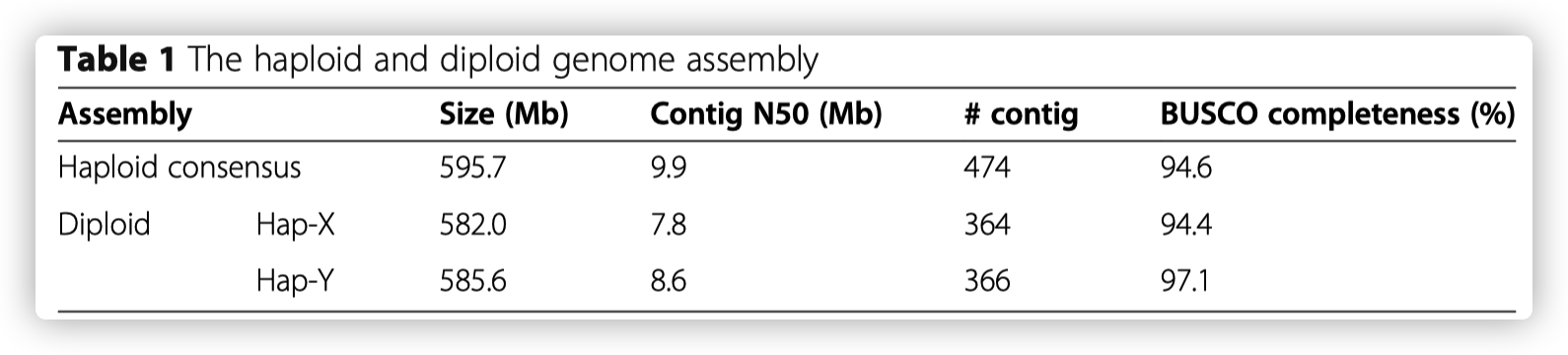
通过提取雄性大刺鳅的肌肉组织DNA进行PacBio三代测序,产生30 G的HiFi数据,结合Hi-C数据进行基因组辅助组装,获得了高质量染色体水平的基因组。基因组大小为600.1 Mb,contig N50为9.9 Mb。组装获得大刺鳅的基因组序列可独立产生两个染色体水平的单倍体基因组hap-X和hap-Y。
首先产生了单倍体一致性contigs,根据它们在现有大刺鳅染色体组装中的锚定位置将其连接到染色体上(fMasArm1.2)。
然后,将HiFi reads比对到染色体水平的基因组上,并结合来自HiFi reads本身和来自同一个体的Hi-C reads的信息进行基因组分型(phasing)。
在hap-X和hap-Y基因组之间,没有检测到大规模的染色体重排,但在新组装和fMasArm1.2组装之间的端粒附近检测到一些倒置。hap-Y基因组组装质量更高,后续分析基于此基因组。
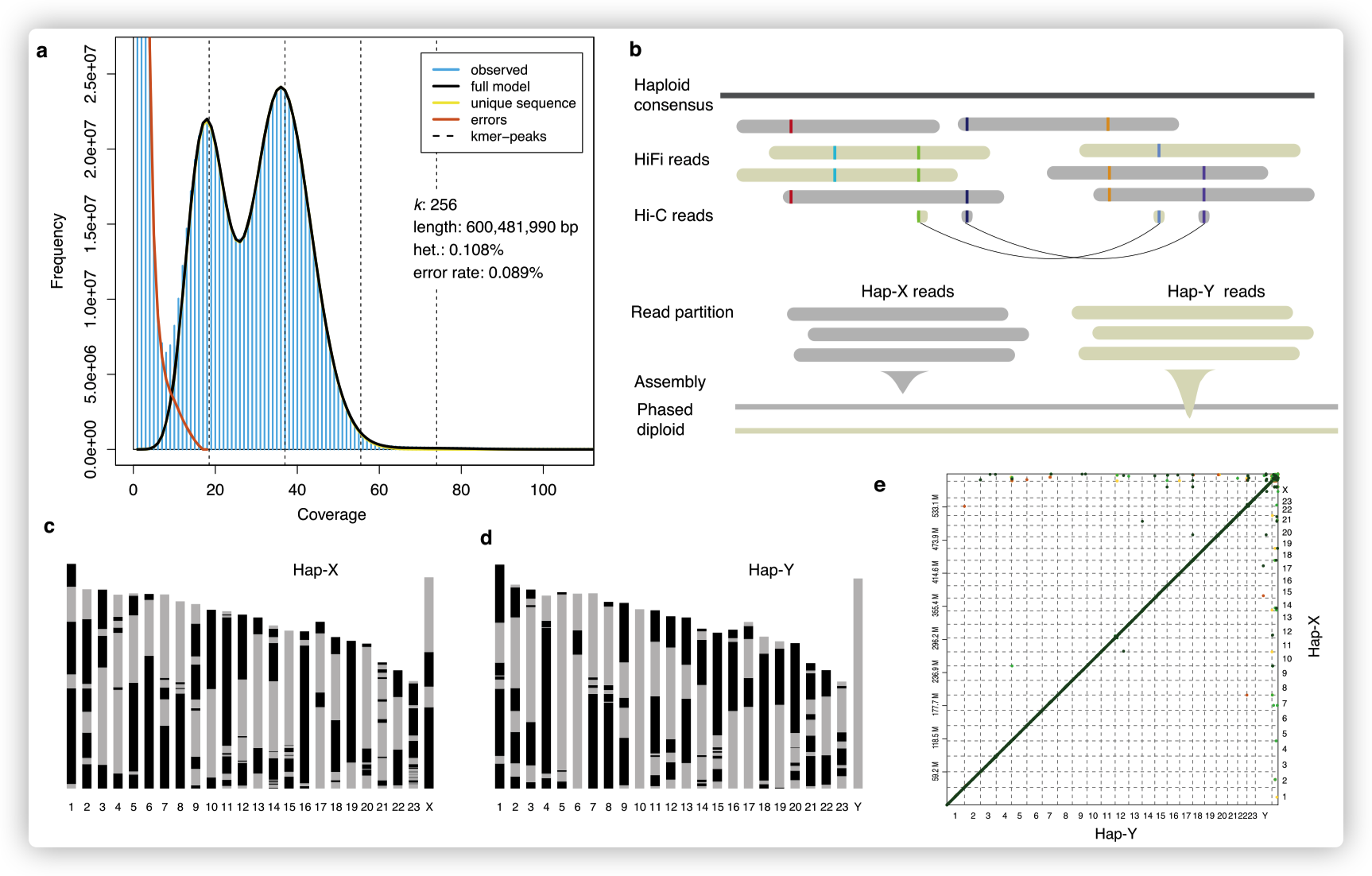
- 将之前鉴定的male特异性标记定位到一个单倍体基因组的一条染色体上,推测该染色体是Y染色体。

- 将分离出的HiFi reads 组装成两个单倍体基因组,它们的基因组大小和contig N50与单倍体一致性组装的基因组相似。
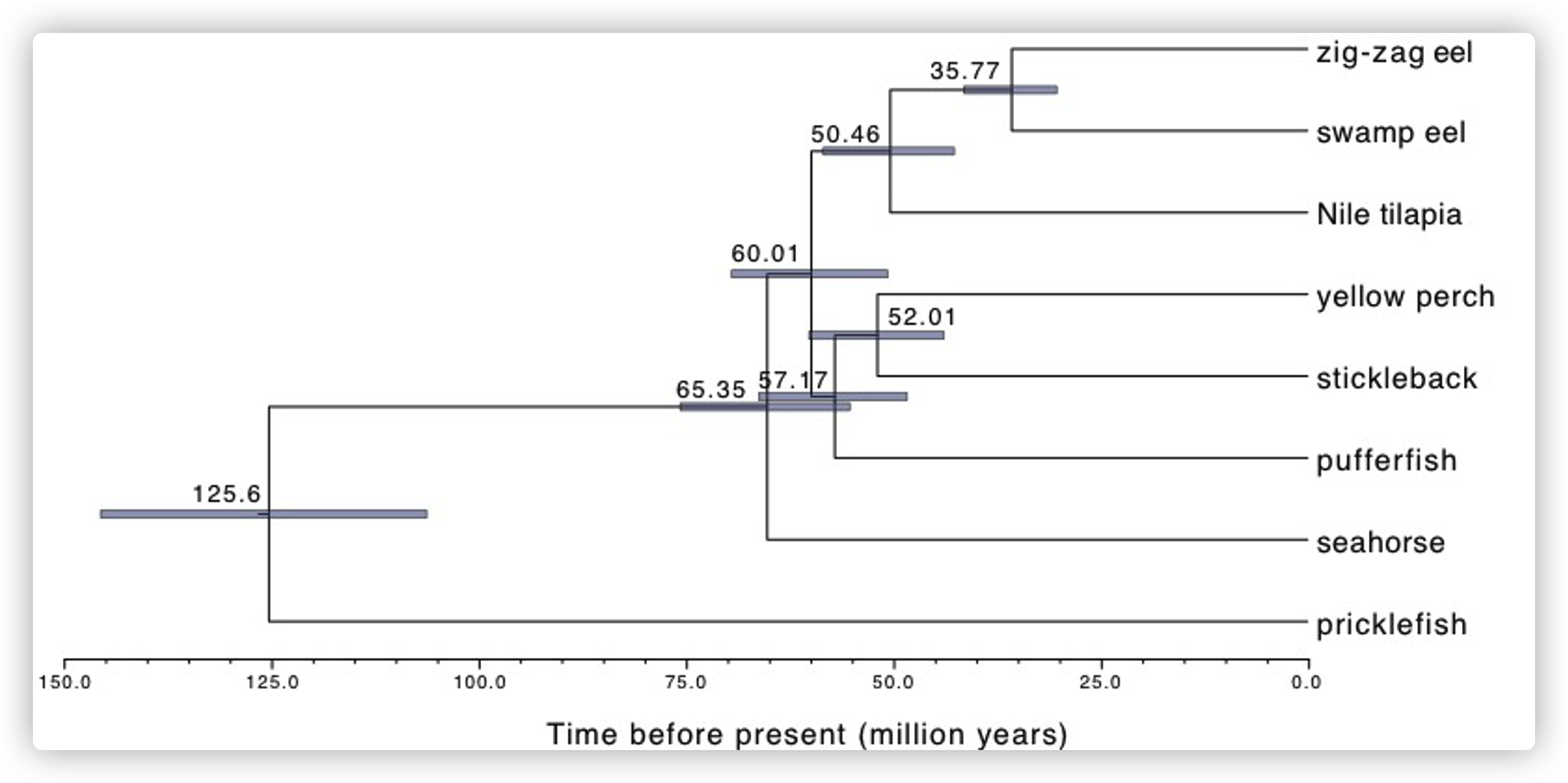
- 通过7个鲈形总目(Percomorpha)物种和一个外群物种Acanthochaenus luetkenii(一种基生棘鳍目鱼类)的全基因组比对数据,确认了大刺鳅与亚洲沼泽鳗有密切的亲缘关系,并估计它们在大约3,600万年前彼此分化。
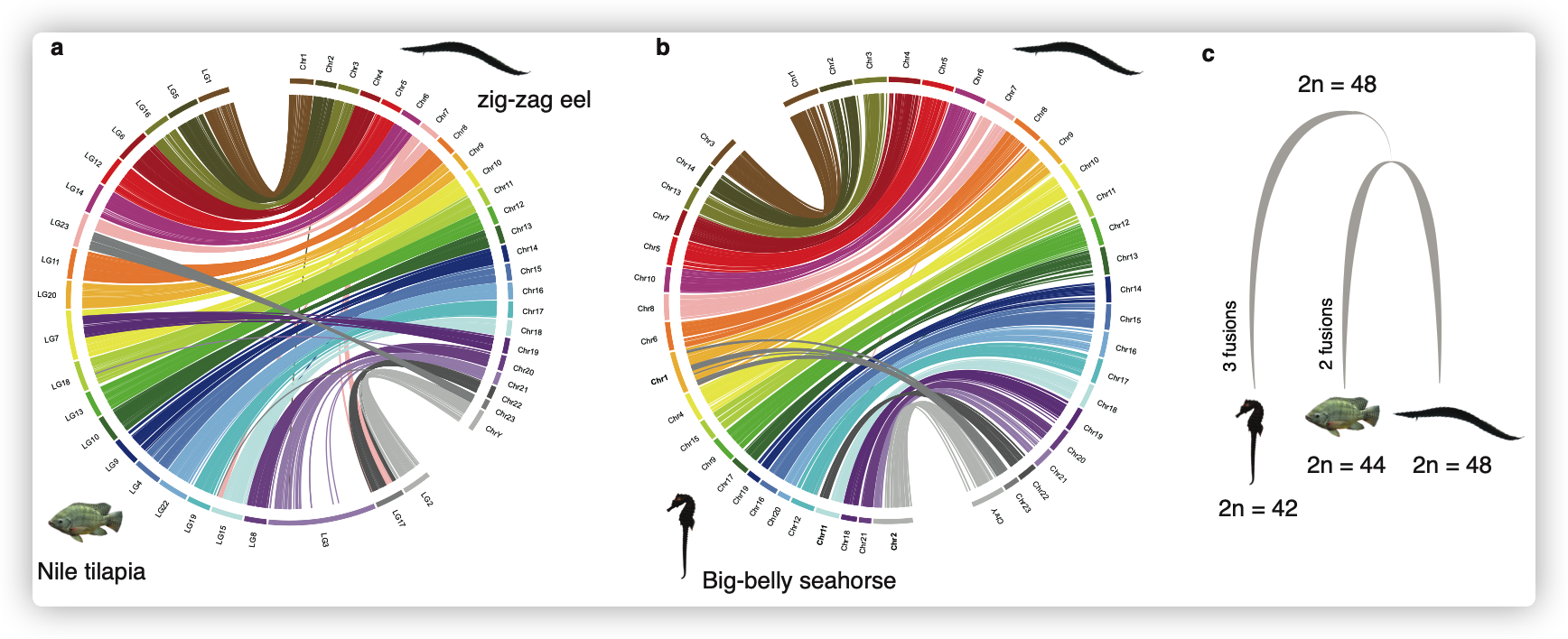
染色体核型进化
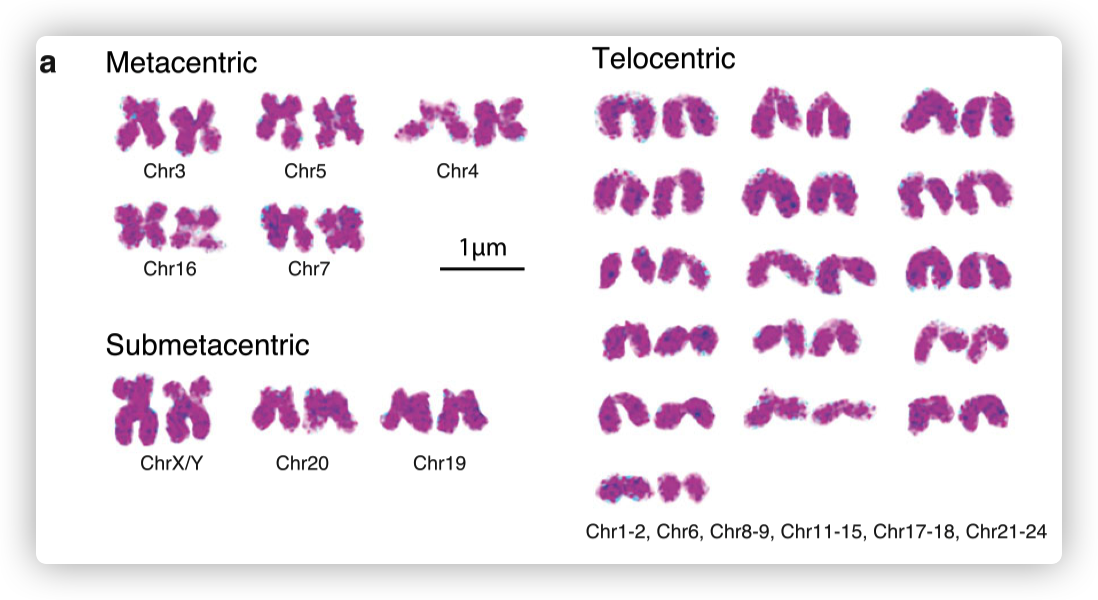
- 根据细胞遗传学观察,将其染色体分为5条中着丝粒染色体、3条亚中着丝粒染色体和16条端着丝粒染色体
- 基因组共线性分析表明,48条染色体是这三个物种(Nile tilapia and big-bellied seahorse)的祖先特征,这与之前的报道一致,即大多数鲈形总目鱼的二倍体数量为48。
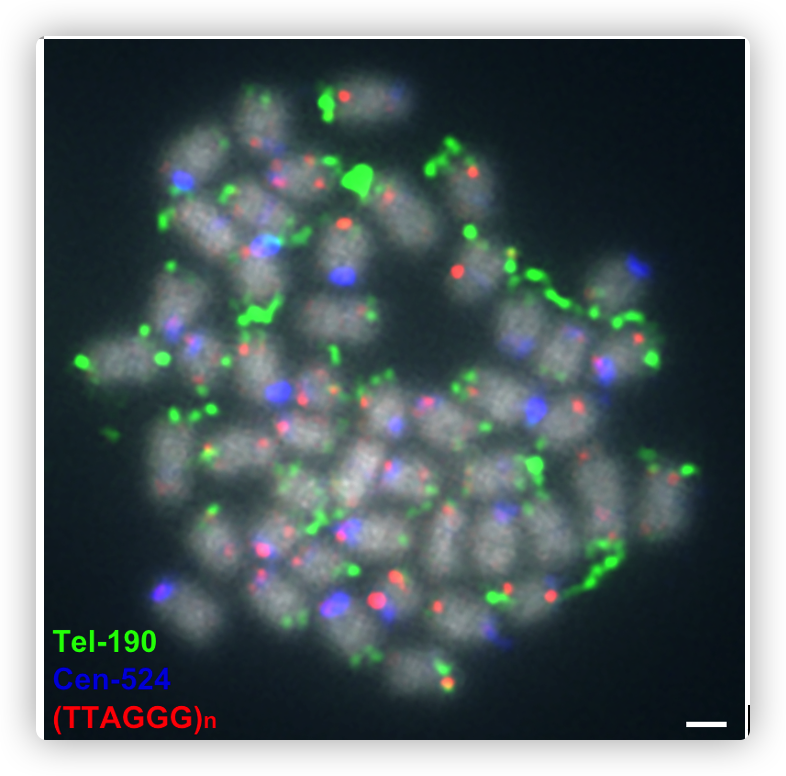
着丝粒卫星序列的基因组和细胞遗传学鉴定
-
为了鉴定着丝粒卫星DNA,根据重复序列注释寻找数量最多的卫星序列。其中两个卫星序列拷贝数较大,单体长度分别为524和190 bp。
-
524 bp的卫星序列(命名为cen524)通常出现在每条染色体上的单个位点,在端着丝粒染色体上,它出现在染色体的一端,而在中和近中着丝粒染色体上,它出现在中间。
Cen-524 a strong candidate for the centromeric satellite -
190-bp的卫星序列(Tel190)只出现在染色体的末端,而在中着丝粒染色体上,有时也出现在两端,这表明Tel190与端粒有关。
Tel-190 is associated with telomeres -
为了进一步验证候选的着丝粒卫星序列,使用荧光原位杂交(FISH)杂交了cen524和Tel-190的探针,发现它们在染色体上的位置与基因组序列组装结果基本一致。

- 大多数着丝粒的组装是不完整的,无法组装大多数染色体的保守端粒序列(TTAGGG)n,可能是因为Tel-190序列非常长,在组装中不完整,我们无法将包含(TTAGGG)n的contigs连接到染色体末端,FISH实验表明(TTAGGG)s序列和Tel-190共同存在。
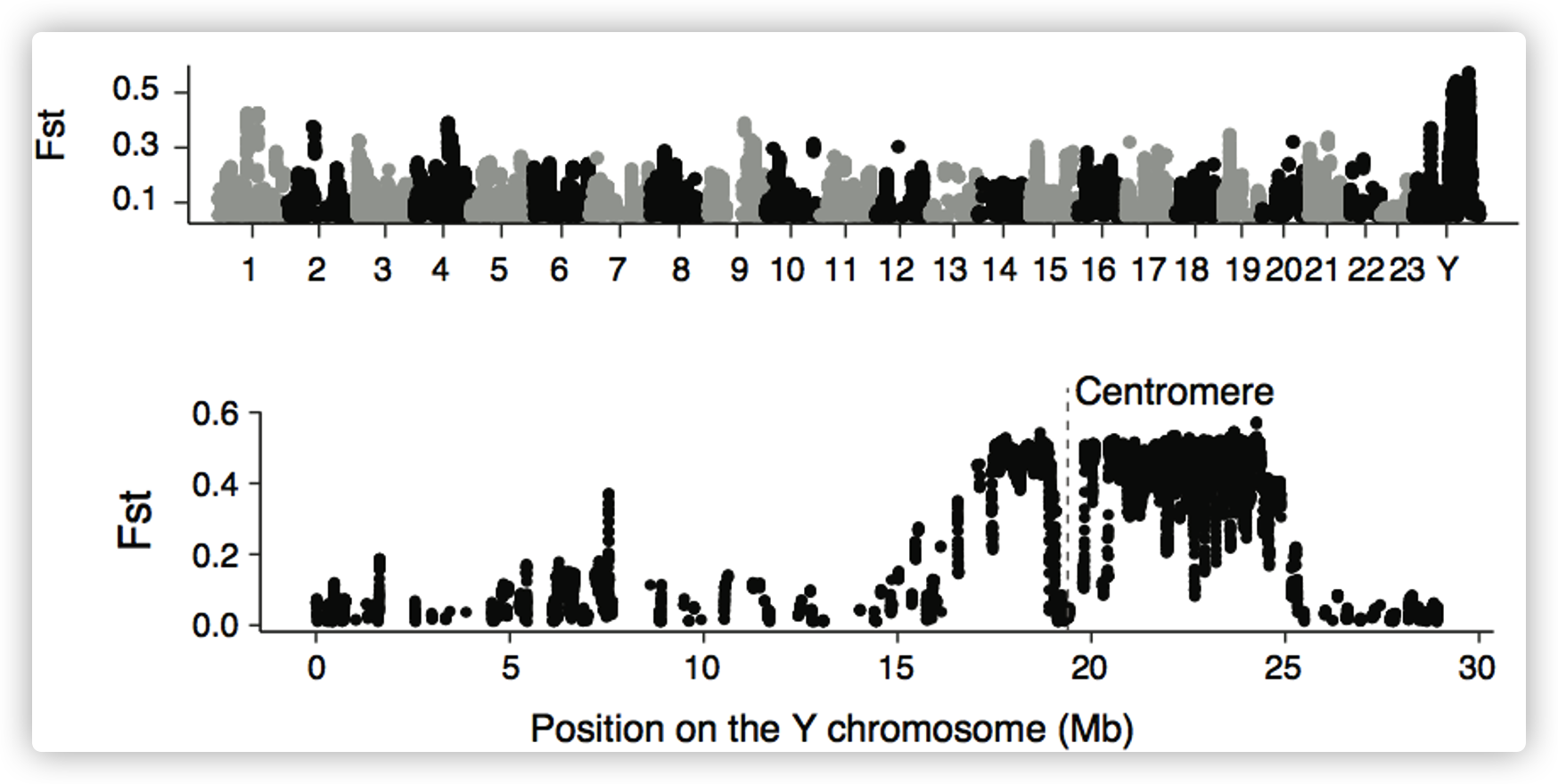
年轻的性染色体
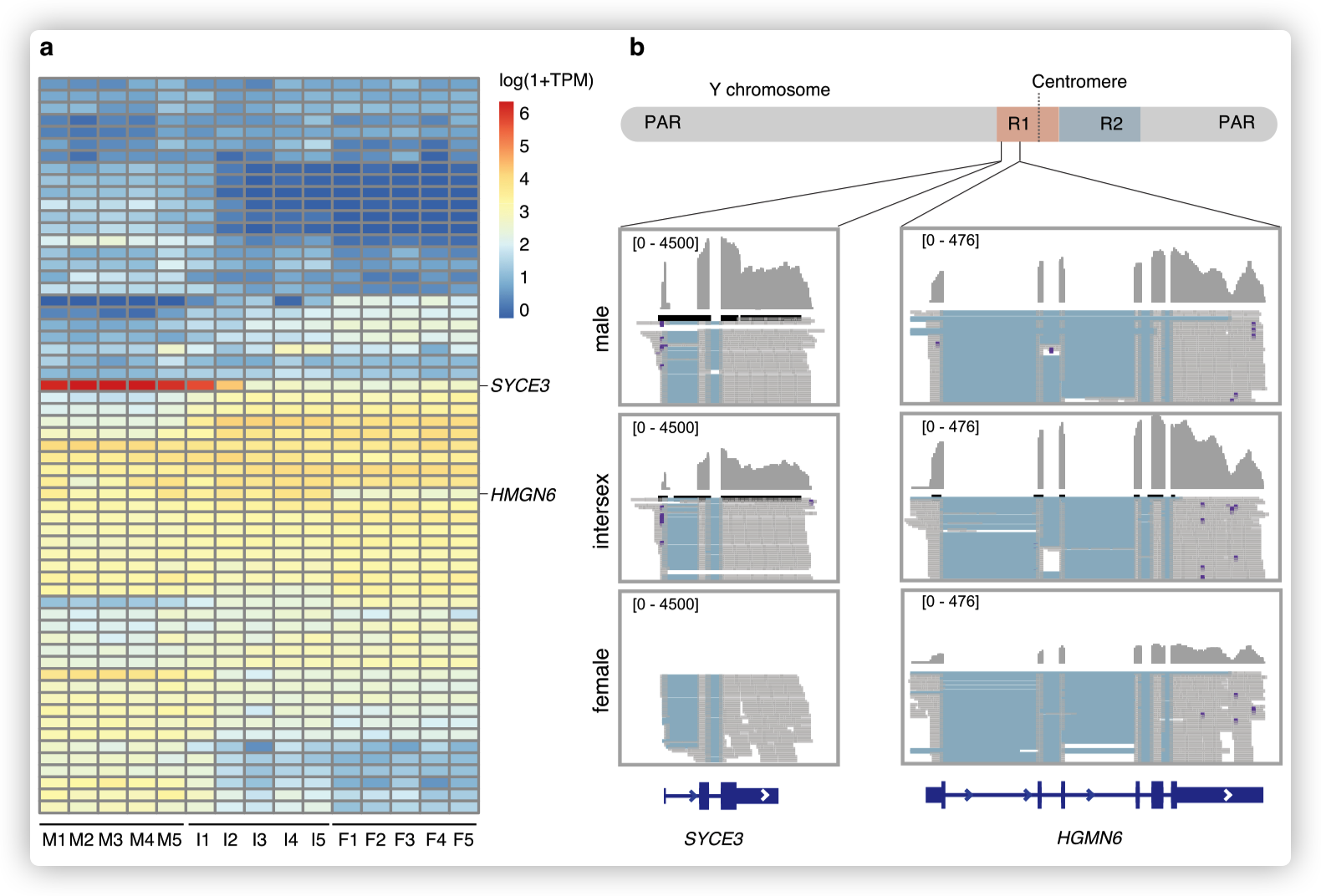
- 对10个male和10个female个体进行了重测序,并筛选了与性别相关的变异。Y染色体上的一个~ 7Mb序列被发现与性别有关,显示出高密度的male特异性变异,与假常染色体区(pseudoautosomal region)或常染色体相比,male和female之间的分歧度(F~ST~)增加。
-
推测SLR跨越着丝粒,染色体的两端为PARs。这表明,在物理上接近着丝粒的位置可能是SLR缺乏重组的原因。
-
第一个区域(R1)具有显著的X-Y差异,并且倾向于比第二个区域(R2)有更高的雄性特异性SNP密度。
-
R1长约3.0 Mb,包含67个蛋白编码基因,横跨着丝粒,而R2基因少,虽然长度长(3.9 Mb),但只包含22个基因。
-
在R1和R2中,X-Y序列的差异接近1%,表明其起源很近,没有检测到这两个区域的染色体倒置或X和Y染色体之间基因含量的差异。
-
R1和R2可能代表了两个进化层次,但我们不能排除只有一个层次的可能性,只是其中一个区域的分化速度比另一个快。
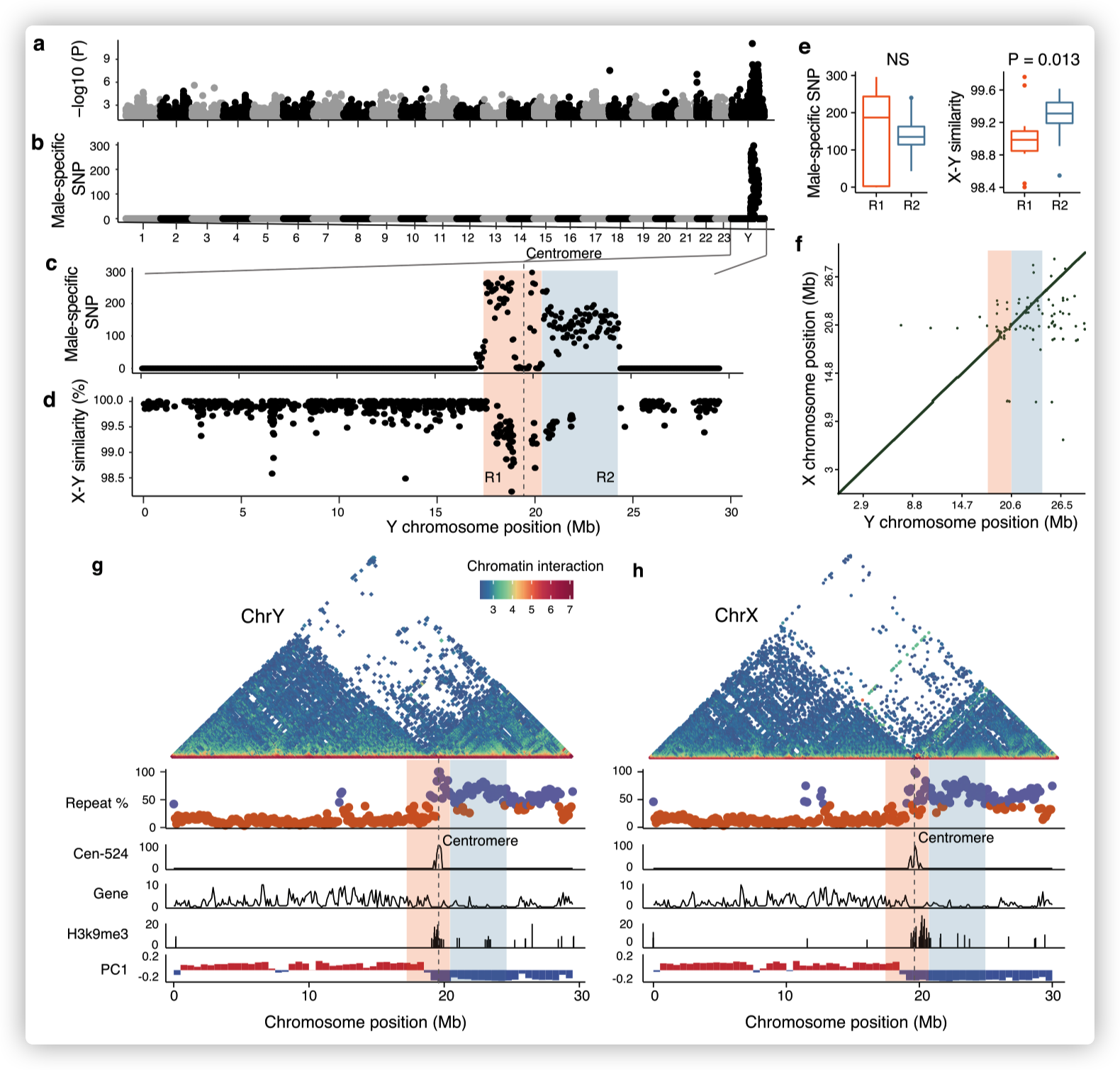
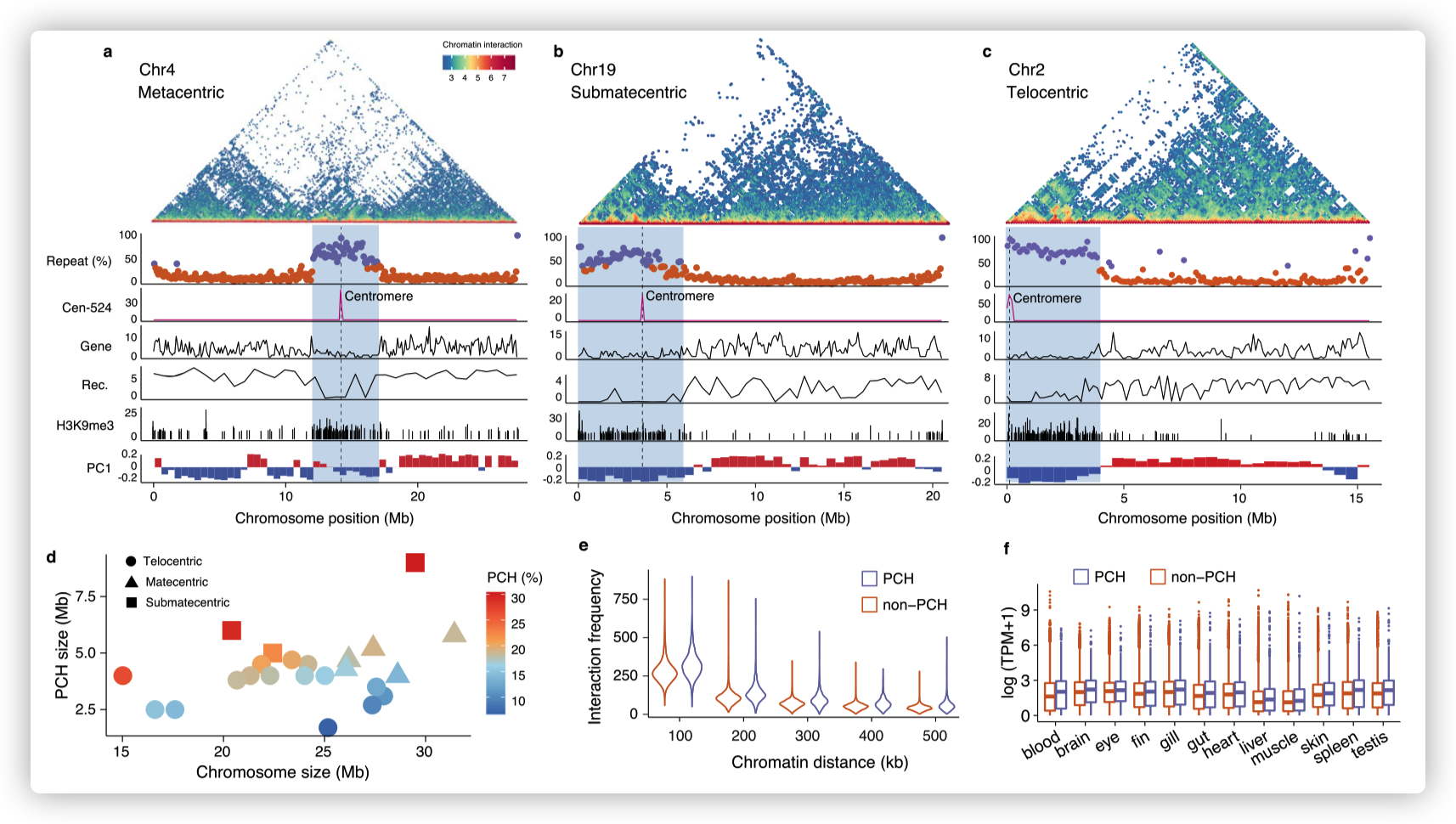
- 大多数的在X和Y染色体的SLR具有高密度的重复序列。整个一对性染色体的短臂,包括X和Y染色体的着丝粒区域,具有典型的异染色质,包括平均重复序列50%以上与长臂(~ 20%),低基因密度,广泛的H3K9me3修饰和广泛的非活性(B)区隔。
The PC1 panel shows the PC1 values of Hi-C epivector: the positive values (red) represent active A compartments and the negative values (blue) represent silenced B compartments.
-
由于其物理位置靠近着丝粒,短臂异染色质可能起源于近着丝粒异染色质(PCH)。广泛分布的异染色质可能是早于X-Y染色体分化的祖先特征,而不是在非重组Y连锁区域进化后衍生出来的,因为它在X和Y染色体上都存在。
pericentromeric heterochromatin (PCH) -
检测了雄性和雌性以及双性个体生殖器官中SLR基因的表达谱,但没有检测到组织特异性表达的趋势。
-
SYCE3,它只在睾丸中表达,但在卵精巢中也有表达(I2和I4)。这表明SYCE3可能参与了精子发生或成熟睾丸的其他生物学过程,比如与减数分裂相关。
-
HMGN6在睾丸和卵精巢早期阶段的表达水平相似,但在卵巢中表达水平低得多,使其成为性别决定的候选基因,可能引导睾丸的发育。
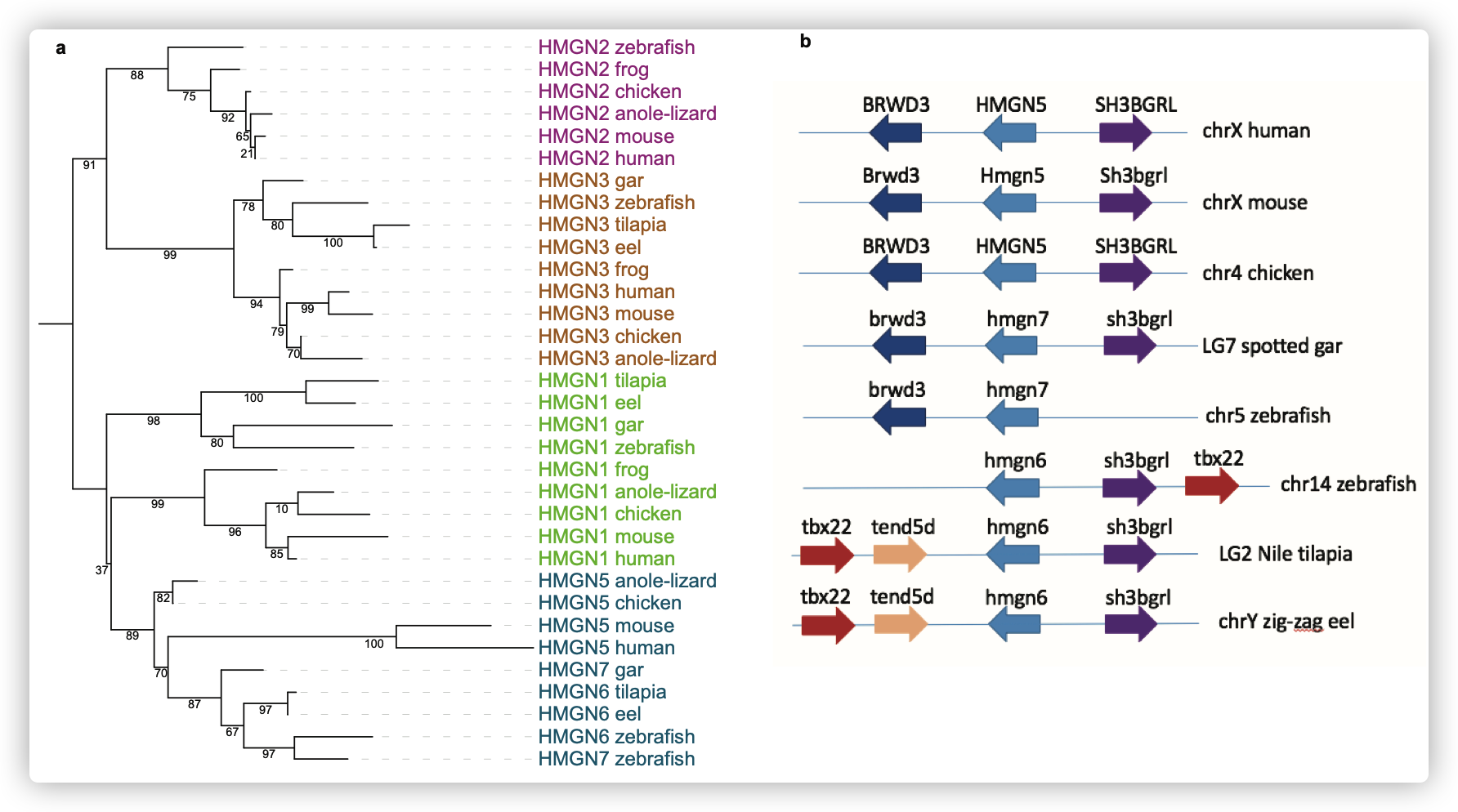
-
HMGN6具有一个HMG (High Mobility Group)结构域,该结构域在哺乳动物主要性别决定基因Sry中也存在。
-
SYCE3和HMGN6都位于R1,SYCE3非常接近SLR和pseudo常染色体区域(PAR)的边界。
- HMGN6似乎只存在于硬骨鱼中,但系统发育分析表明它与HMGN5密切相关。
广泛的近着丝粒异染色质
-
着丝粒周围较大区域(~ 4mb)的重复含量较高,通常高于50%,而基因组其余部分的重复含量低于16%。这些高度重复区域具有较低的基因密度、较低的重组率和更频繁的H3k9me3修饰,与PCH一致。
-
较小的染色体,特别是端着丝粒和近端着丝粒染色体,具有较大的PCH比例。这包括XY染色体,它们是近中着丝粒的,几乎整个近中着丝粒染色体的短臂都是异染色质的。
-
在着丝粒周围区域,染色质在较大物理距离下的相互作用更频繁,这与它们较高的折叠和压缩程度一致。
-
发现与其他区域相比,近着丝粒区域高表达水平和高表达宽度的基因比例更大,这与之前的观点一致,即H3K9me3在基因抑制中发挥有限的作用,可能存在其他表观遗传修饰调控PCH中基因的表达。
讨论
-
这是二倍体基因组中唯一的无gap染色体(Y 染色体),X-和Y-linked区域之间的差异虽然低,但可以更清晰地划分单倍体特异性reads,帮助解决重复区域。
-
高质量基因组显示,近着丝粒区域可以占据相当大的比例(6.8至30.6%)的染色体,PCH对鱼类三维基因组的构成的影响是未来的研究课题。
-
蓝罗非鱼的Z染色体上的SLR区域是高度重复和异染色质化的。
-
鉴于大刺鳅的选育品系需要4年才能达到生殖成熟,对候选性别决定基因的功能验证是具有挑战性的。
-
然而,我们对雄性、雌性和双性个体生殖器官基因表达的分析表明,HMGN6是决定性别的一个强有力的候选基因,它在雌雄同体时时表达。
-
许多与性别决定相关的转录因子都含有HMG结构域,包括SOX9和SRY。
参考
https://doi.org/10.1186/s13059-021-02430-y
Perspective
- Sex chromosomes can evolve once recombination is halted between a homologous pair of chromosomes.
Why do sex chromosomes stop recombining
性染色体进化的性拮抗模型
- recombination will be selected against in the region between a sex-determining gene and a nearby gene with sex-specific effects
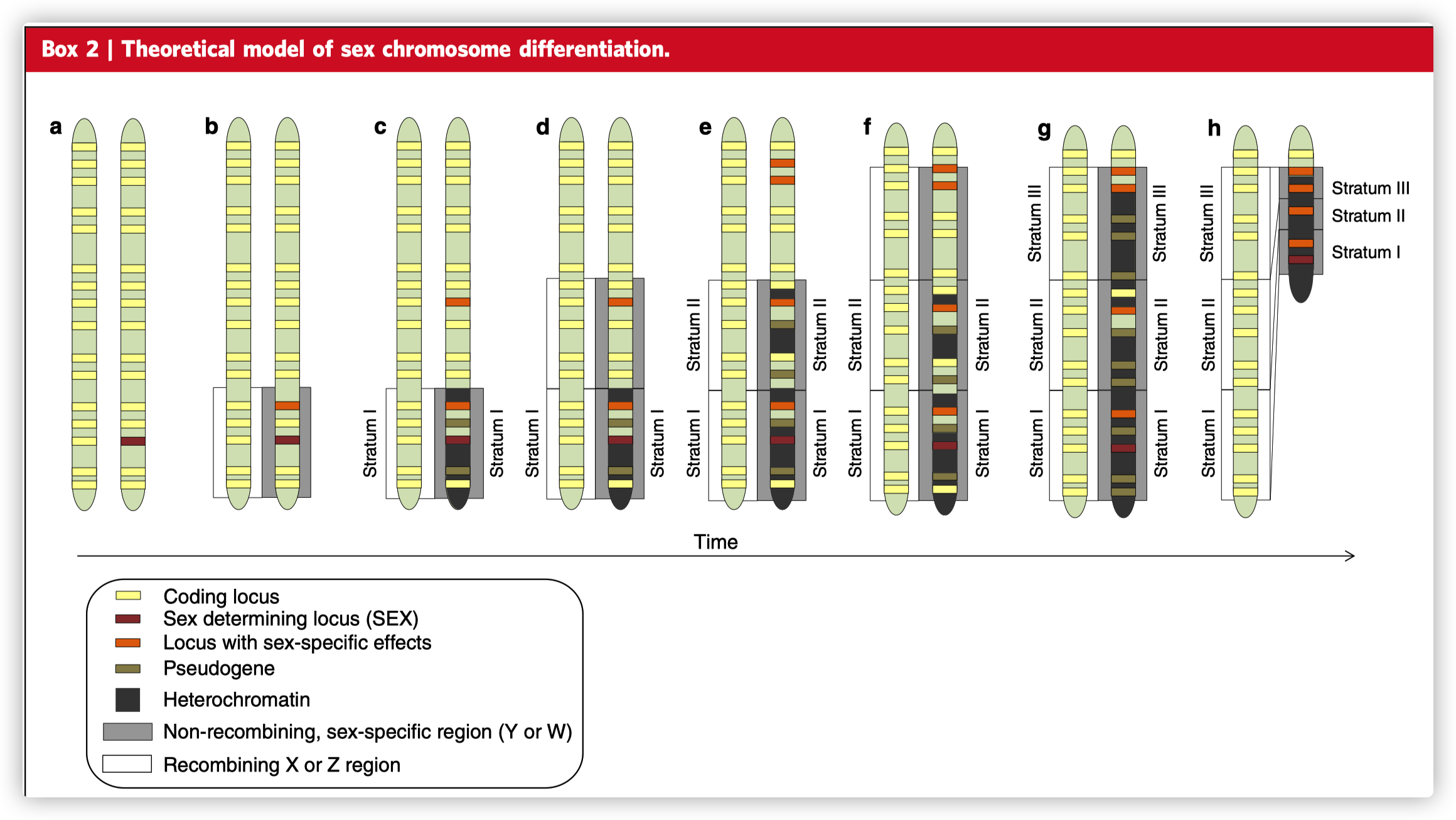
性染色体从常染色体进化而来,最初是获得了一个性别决定位点(a)。在靠近性别决定位点的位点上出现的性拮抗等位基因导致X和Y或Z和W染色体之间的重组抑制(b),产生第一层(Stratum I),它逐渐异染色质化。一旦Y 或 W染色体上的重组被终止,没有性别特异性的基因往往被假基因化。不重组区域可以随着额外的性拮抗等位基因的获得和进一步的重组抑制而扩大,导致额外的X-Y或Z-W strata—spatial clusters,具有类似的分歧时间,在哺乳动物、鸟类、鱼类和植物中观察到,它们也经历了基因功能丧失和异染色质化(d-g)。重组的缺乏导致重复性DNA序列的积累,这可能导致Y或W的大小短期内增加,但通常会导致大规模的缺失,性染色体的物理大小大量减少,以及产生高度异形的性染色体(h)。
- Stratum: Region on the sex chromosomes where recombination has been suppressed. Strata can be identified by spatial clusters of X-Y or Z-W orthologs with similar divergence estimates.
从雌雄同体到性染色体的转变
在植物和其他系统中,性染色体与从雌雄同体到分离性别的过渡有关,性染色体的形成可能采取与祖先分离性别的物种略有不同的路线。在这种情况下,显性模型预测,同一染色体上单独的雄性和雌性突变通过雌雄同体的中间阶段导致从雌雄同体到两性的转变。一旦这些突变发生并在种群中达到足够的频率,它们之间的重组抑制就会防止逆转为雌雄同体,导致性染色体的进化。
How is recombination halted between the sex chromosomes
跨越性别决定位点和附近的性拮抗位点的染色体倒位通常被认为是停止了重组,因此推动了性染色体的分化。有间接的证据表明倒位与性染色体的进化有关系。例如,许多动物和植物的性染色体显示了Stratum的证据,即具有相似分化估计的X-Y或Z-W,这些空间的cluster与染色体倒位事件瞬间停止了所有包含的基因座的重组是一致的。
Why do sex chromosomes diverge at such different rates
同态性染色体的普遍性
- 许多具有遗传性别决定的生物体缺乏异型性染色体,这表明非重组区域尚未在性别决定位点以外显著扩散。
Age
- 一些同态性染色体是年轻的,可能处于退化的早期阶段,年龄并非总能准确预测非重组区域的相对大小,因此也不能准确预测整个性染色体的差异。
Relative length of haploid phase
- 有些生物体具有较长的单倍体时期,从而产生较强的单倍体纯化选择,以维持Y染色体上的基因活性。
Sex chromosome dosage compensation
- 这种退化的结果是,相对于异配性别的常染色体,X和Z染色体上的基因剂量减少。这种基因表达的不平衡通常被认为是有害的,并扰乱相互作用的基因产物的生化计量。这些有害影响被假设为驱动剂量补偿机制的进化,以恢复祖先的二倍体表达水平。剂量补偿的程度某些类群中差异显著,尽管一些物种表现出完全和不完全的性染色体剂量补偿效应。
Sex reversal
- 性别逆转,即个体表型和基因型性别之间的不一致,可能在重组抑制和性染色体进化中起重要作用。
Sexual conflict
- 性拮抗的等位基因是性染色体进化的性拮抗模型的核心。有性拮抗的非重组区域区的快速扩张是因为在基因组内拥有更多位点,进而更加靠近性别决定位点,并携带性拮抗的等位基因。